


| Sponsors partenaires du MCF, informations et conditions par mail : CONTACT | |||||||
|
|
|
|
|
||||
SOME GENETIC CHARACTERISTICS OF THE MARANSTranslated from the French by Katherine Anderson (Marans Club USA). |
|
|
![]()
GENETIC CHARACTERISTICS OF SHANK FEATHERING |
The presence of the trait of feathered legs is crucial in the Marans. It constitutes one of the essential marks of the breed.
This characteristic was introduced into the Marans through crossings with Asian breeds of the Langshan type. It is also present in many breeds, but it is, expressed in different ways according to the type and the combination of modifying genes present in the genotype considered.
The symbol of the presence of “feathered shanks” is expressed by (Pti), an autosomal dominant gene, however, it can comprise three different types according to its importance in the feathering of the various parts on the legs of certain breeds.
In regards to Marans, only the lateral sides of the shanks as well as the two toes on the lateral portion of the feet are feathered, and that always in a reasonable proportion. Indeed, from the point of view of its extent, sparse feathering is of course desired and when it is a little too invasive especially in the presence of “cuffs”, this in particular betrays a return of traits from the original crossings with the Brahma, Langshan.
In any event, this trait of feathered shanks and lateral toes in the Marans constitute a rather easily selected characteristic.
When the subjects show a clear absence of any feathering on the shanks, they must be eliminated from a breeding programme because they can engender a loss of the obligatory dominant (Pti) gene.
The showing of such individuals as English standard Marans does not rule out the problem.The true Marans is none other than the original Marans comprising all the integrality of its genes, “feathered shanks” included.
GENETIC
CHARACTERISTICS OF
|
In general the genetic characteristics responsible for the normal and fast growth of the plumage in poultry is recessive and sex-linked. It is symbolized by the term (k+). So that the subjects in question sport normal plumage and not slowed growth (presence of k+), it is imperative that three dominant factors of the series K are missing from their genotype, which are: (K), (Ks) and (Kn), called “Asian slow-feathering” genes.
In addition, poultry affected by the gene (k+) will be really normal for plumage growth only if carrying, moreover, the dominant factor (T+).
In the absence of (T+), the subjects (k+) will thus be affected by the presence of one of the recessive factors of the series T, which are (t) or (ts), called “autosomal slow-feathering” genes.
In a way, a chick whose appearance and growth of the plumage are naturally fast will thus genetically consist of the unit [(k+), (T+)].
The wild breeds of the Bankiva type as well as many other breeds of the Mediterranean type have fast feathering, which upon hatching have 6 already emerged primary flight feathers and almost all the secondaries quite visible; they are thus [(k+), (T+)].
(k+) for: absence of Asian slow-feathering
(T+) for: absence of autosomal slow-feathering.
We have incontestably detected the presence of these types of reducing genetic factors in certain lines of Marans, in particular genes (K) or (Ks), or even (t) or (ts).
Those are, on the other hand, not easily identifiable between them and subject to complementary experiments in the breedings followed by the MCF.
This characteristic can be easily located by the presence of slow or late fledging of chicks until at least 10 days and even more according to some cases.One notices only a few or no feathers on the tail, flight feathers, and on the back.
Let us quote this after some elements of recognition of these various types of slow fledging in order to facilitate their possible location in young subjects.
GENETIC CHARACTERISTICS OF THE TYPE (K) ASIAN SLOW-FEATHERING GENE
Factors concerning (K), (Ks), and (kn) influence the speed of starting and growth of the feathers on various levels, actng as a “brake” in fast-fledging chickens in the very first weeks of age.
-1- the gene (K):
It simply causes late feathering in some subjects and it is present in several well known heavy breeds, generally of Asian, American or English origin, such as the Brahma, the Indian Game, the Cochin and theWyandotte. One understands that the Marans can be occasionally affected by this type of genetic trait inherited from some of its Asian ancestors.
-2- the gene (Ks):
It generally causes slow or very slow fledging during the first 12 weeks of life; it is then without any influence in adulthood. Some Marans could also be affected with this trait.
-3- the gene (Kn):
This gene causes very slow fledging, almost truly handicapping insofar as one can note a true limitation of growth of the plumage throughout almost the whole of its juvenile development and even into adulthood. It can still cause a limitation of the size of the comb as well as an atrophy of the uropygial gland.
Marans do not seem affected by this last allele, but if necessary, it must be radically eliminated.
GENETIC CHARACTERISTICS OF THE TYPE T AUTOSOMAL SLOW-FEATHERING GENE
As the inverse of the Asian slow-feathering gene (k) examined previously, there are still two different factors highlighted in 1933 and then in 1946, of the types (t) or (ts) which are also responsible for a real slowing of fledging.
As for the animals known as normal, they are carrying the dominant autosomal gene (T+), i.e. non-sex-linked and which acts only in the presence of (k+) in the genotype: i.e. the absence of any Asian slow-feathering gene.
-1- the gene (t):
It causes a slow fledging, but a little less late, i.e. until the age of six weeks maximum. Marans could be affected with this type of allele.
-2- the gene (ts):
This gene causes definitely late feathering, at least until the eighth week of age. Marans could also be occasionally affected, but this still merits confirmation.
REVIEW OF MARANS AFFECTED WITH SLOW-FEATHERING GENES
Based on the state of the experiments to date and those which deserve further study, we can summarize the phenotypic differences with the following elements:
At hatch:
The normal chicks [(k+), (T+)] do not have any slowing of feathering have their primary and secondary flight feathers and visible wing coverts.
The flight feathers are more developed than the coverts.
Conversely, the chicks affected with slow-feathering genes do not show longer flight feathers than the coverts.
At ten days:
The difference between the various genotypes is clearer than at hatching.
The normal subjects [(k+), (T+)] must have a tail of approximately two centimeters and their flight feathers almost cover their sides to the tail.
The subjects (K), on the other hand, do not have a tail and their flight feathers are already well-developed.
The subjects [(k+), (ts)] do not have a tail and their secondary flight feathers are normal, but there are only three or four primary flight feathers.
The subjects [(k+), (t)] have a tail and there are six very shortened primary flight feathers and no secondary flight feathers, or at most, very few.
At eight weeks:
The normal subjects [(k+), (T+)] show a tail which has started to moult from the center.
On the other hand, the subjects with “slow-feathering” genes have frayed caudal feathers, which are softer at the base.
PROBLEMS CAUSED BY (K) OR (t) SLOW-FEATHERING GENES
It is advisable to remember here that once adulthood, i.e. sexual maturity, is reached there is no difference in plumage between the various subjects whether or not they are affected by this phenomenon. Sorting between them becomes impossible after this age and often even much earlier.
We have to thus make a type of selection made easier at hatching. Simply eliminating the young chicks locatable by difficult fledging, in particular on the tail, the back, and the wings would make it possible to obtain a rather easy cleansing of the stock over a few years.
In addition, since during expositions or shows, it is not possible to distinguish the subjects concerned as they are adults, then we are very tempted to think that these types of reducing genes do not present major disadvantages. In fact, the risk exists of a real and lasting laxity in regards to selection for breeders from this point.
It appears then that these types of “slow-feathering” genes of the young chicks, which are of course hereditary, not only do not bring anything positive to the breed but especially can cause some problems.
Thus, it was discovered in 1988 that the Asian slow-feathering gene (K) in particular would be in close connection with the existence of an endoviral gene of the viral family which causes leucoses. The subjects with the slow-feathering gene (K) would be thus more sensitive to this group of diseases and have proven to have reduced performance.
In addition, slow or late feathering makes it obvious that these young subjects are more sensitive to picking, having their back stripped to at least four weeks of age and especially, their puny tail feathers which are not very hard at the base until at least eight weeks of age.
Let us recall that at this age, the normal subjects have their caudal feathers entirely finished.
We must still stress that the subjects with “slow-feathering” genes show after slaughter a more significant number of “pinfeathers” than normal subjects.
Lastly, we must add that the young Marans pullets with normal and rapid feathering tend to begin laying earlier. To contrast, allele (K) might, however, make it possible to support a greater clarity of “barring” in the Cuckoo plumage, or even of the intensity of the red in poultry plumage.
In spite of the probability of this attraction, which remains to be shown at a greater length, these types of genes are proving more or less handicapping and rather not easily identifiable between them. Consequently, it seems quite obvious, as far as possible, to limit if not eliminate all the young Marans subjects which present one or another of these reducing genes.
|
THE GENETIC CHARACTERISTICS OF VULTURE HOCKS OR CUFFS |
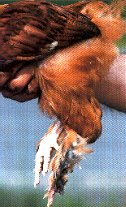 |
The cuffs, also called “vulture hocks” or “vulture boots” consist of very lengthened feathers located on the back face of the thighs moving towards the rear of the animal. Moreover, they are always associatied with the presence of “feathered shanks.”
The cuffs are due to a genetic autosomal and recessive factor which is symbolized by (vh).
We can identify them as present and desirable in certain breeds such as the Sabelpoot, Sultan, Breda, or Bearded d’Uccle…on the other hand, in the Marans like the majority of the feather-legged breeds, it is a serious defect to be eliminated. This defect is however not very frequent in the Marans.
These long and rather stiff feathers extend behind the thighs and beyond the articulations of the shanks. This anomaly appears in particular among the principally recalcitrant defects to be eliminated within the Brahma breed, and that can undoubtedly explain the existence of its occasional atavistic return in the Marans.
Marans dressed out in such cuffs must be, of course, avoided when selecting for breeding without the least hesitation.
It is thus advisable to avoid spreading such
an imperfection any more than possible as here again we have a
recessive gene, and it is to say theoretically irreversible if it
becomes widespread.
|
GENETIC CHARACTERISTICS OF SPLIT WINGS |
 |
Split wings constitute a serious hereditary anatomical anomaly in all poultry breeds. When the wing is opened, they show a more or less significant space between the group of primary flight feathers and the group of secondary flight feathers.
These two groups of quills must always join tightly in a complete wing in order to ensure effective flight.
This imperfection of split wings is due to an anatomical defect in the construction of the skeleton and tendons of the wing and appears more often in the light breeds.
According to Brandt and Willems, this trait of split wings would be a simple autosomal recessive genetic factor, and today still not symbolized.
In addition, it is advisable to specify that other quite as serious closely related anomalies exist, namely, flight feathers curled in a corkscrew, quills which cross each other, or sagging wings. We noticed that the defect of split wings in certain poultry was concomitant with a reduction in the width of the flight feathers. Thus, it was noticed that when the flight quills present themselves as narrower and more frayed in addition to having separated barbules, these signs resemble other defects than split wings; as in this example. In the Marans, split wings constitute a frighteningly tough hereditary defect which one can control only by strictly followed vigilance in eliminating all recessive subjects from the breeding programme.
|
GENETIC CHARACTERISTICS OF HARD PLUMAGE AND CLOSE-FEATHERING OF THE BODY |
Many breeds of fighting games have hard plumage laid close to the body, caused by the genotypic presence of a dominant allele symbolized by (Ha).
Conversely, the other type of plumage is puffed out or loose, for example, which one finds in Cochins or Orpingtons, and is a recessive factor expressed by (ha+).
This gene should never relate to the Marans breed, as soft and puffed out plumage is prohibited.
This aspect of more or less close plumage is of particular importance in the Marans. Indeed, this is likely to directly influence the quality of presentation of the typical general silhouette of the Marans.
That being known, it should however be recognized that this type of hard plumage or soft plumage seems also fed by a minimum of complexity, though not elucidated.
Indeed, there are rather numerous cases of breeds presenting a more or less intermediate feathering between these two extremes expressed above, we are logically brought to suspect in this respect the possible existence of complementary genes which would come to affect the expression of alleles (Ha) and (ha+).
Though it exists, Marans should never be tolerated with puffed out plumage.
Thus, contrary to other breeds, when one seizes in one’s hands a good Marans hen one is often surprised by her real mass that her appearance did not translate. In other words, the Marans always comprises a minimum of tight plumage which tends to hide its mass.
|
GENETIC CHARACTERISTICS OF SHANK COLOURATION |
The skin colouration of poultry (for example, white skin or yellow skin) and the dermis of the shanks can be independently affected by the presence of specific genes responsible for leg colour in poultry.
There are indeed two different genes expressed by the signs (id+) and (ID) which are responsible for the presence or absence of “melanin” (black pigments) in the skin of the shanks.
The gene (id+) imposes the appearance of melanin in the dermal layer of the shanks, which then shows a bluish gray color in chickens with white skin (in the case of the white Bresse, for example).
However, found in the genotype of a breed with yellow skin, the shanks then appear olive-green (black breasted red English Game and certain Northern Games).
This genetic darkening characteristic of the legs (id+) is a recessive sex-linked trait.
The dominant gene (ID) causes, on the contrary, the absence of melanin on the level of the shanks, which are then expressed as rosy-white in the breeds with white skin (Marans or Gâtinaise, for example) or yellow in the breeds with yellow skin (Northern Games, Wyandottes).
On this subject, let us benefit from the occasion to recall that certain genes for plumage colour like “cuckoo” (B), “wheaten” (Ewh), or “splash” (Bl/Bl) are accompanied by a hereditary inhibition of the melanin, which explains in particular why all poultry breeds with “cuckoo” and “wheaten” plumage as in the Marans, have invariably clear shanks beaks and nails which are of course genetically dependent for their colour.
Nevertheless, for the Marans varieties with dominant black genes such as the self-coloured black, Silvered Black [Birchen], and Black-copper [Brown-red] the presence clear shanks involves an additional difficulty in the selection of breeding stock. In effect, this trait often appears in conjunction with the presence of light or white down or undercolour along with the appearance of unwanted white feathers at the base of the tail or wheaten flight feathers if they are not totally white.
|
GENETIC CHARACTERISTICS OF OFF-WHITE EARLOBES |
The presence of off-white earlobes is a sign of genetic impurity by improper selection or atavistic expression of the factor for “white earlobes” in the descendants.
It is advisable to note here that the colour of the earlobes, red as well as white, should be evaluated only when the subjects reach adulthood.
The genetic factors responsible for the white or red colour of the earlobes are due to a whole complex of associated genes which are to date unknown; the symbol remains unspecified.
Marans with white earlobes, or more simply off-white are of course not accepted as standard. They should have, as for about half of the other French breeds, only red earlobes without defect.
It should be noted that this likely resulted from the contributions of the blood of Asian breeds which were imported at the end of the 19th century, whereas the majority of breeds with white earlobes are descended essentially from the very old European and Mediterranean breeds.
|
GENETIC CHARACTERISTICS OF SPURRED HENS |
The presence of spurs in hens after sexual maturity is a rare phenomenon today and is of course considered a serious defect for Marans as in the majority of our poultry breeds.
This characteristic was described in 1931 as being hereditary and consequently of genetic origin. In addition, it appears that as of the 17th century, there were already rather frequent cases of hens with spurs, which did not have a great reputation because of the brittleness of their eggs.
According to experiments which have been undertaken since approximately 1925, the hereditary genes involved appear very complex and they overlap those which seem recessive and others which would appear partially or completely dominant.
The heredity of the genetic trait remains unknown today. Thus the symbol of this allele was never defined.
It is advisable to report here the fact, already extremely old, that in regard to the true pit-fighting breeds, the spurred hens were carefully selected for with an aim of transmitting increased length of spurs to the fighting cocks. They bring, moreover, one proven benefit of strength and vitality for the hens and their descendants.
Attention must be paid, however, not to confuse genetically spurred hens with those often older hens which because of disease, accidents or wounds see the cessation of sexual activity and the unbalancing of their hormonal proportions. Such hens carrying ovarian anomalies can then comprise a reduction or even a disappearance of any female hormonal secretion and the subsequent development of spurs.
This phenomenon of accidental origin does not, in concrete cases, have anything to do with the genotypic presence of the factor for “spurred hens” described above. Obviously, one may easily understand that neither genetically based individuals nor the others are exploitable for correct breeding.
|
GENETIC CHARACTERISTICS OF THE FIGHTING COCK |
 |
The characteristics of combat in certain breeds known as “fighting games” are due, not only with components of hereditary origin specific to the breeding drive, but also components of purely genetic origin.
The genetic factors responsible for this faculty or will to fight are not identified to date. It seems indeed, that according to experiments carried out with fighting games, the hereditary factors responsible involve a very complex transmission. In addition, it was noted that the cocks had a greater hereditary influence in regards to transmission of the two various characteristics or styles of combat, which are endurance and speed. It is thus impossible to deny a purely genetic origin of the aptitude for the combat of the fighting breeds.
In regard to Marans, the competition between cocks is always severe, particularly of a chivalrous and enduring type, but it is still possible to note an aptitude for combat between two hens for reasons of social competition. These fights are always very short, but their style is rigorously the same as in engagements between cocks.
From this point of view, it is possible to admit that a genetic influence resulting from the Marans’s fighting game ancestors can remain today in limited proportions.
When certain cocks are vigorous and particularly aggressive towards people or even the stockbreeder, they are often forsaken or even eliminated from many breeding programmes to the detriment of the superior vitality that they naturally exert in their social environment.
This selective sorting appears particularly harmful to the benefit of good fecundity in the hens and a possible improvement in the hardiness of the stock, which we have the right to hope for on behalf of the cocks.
|
HEREDITY OF PLUMAGE COLOUR IN THE MARANS |
 |
|
The MCF publishes a booklet explaining the work of certain crossings between different plumage colours in the Marans with the possibilities of obtaining new stock. It is signed by Albert Roguet, Vice-President of the MCF. This work on the genetics of plumage colour is in the process of continual improvement. This booklet is available near the head office of the MCF for the price of 7 euros (French language). |
Without entering into detail of the origin of chicken breeds, we can say that the great diversity of the plumage colours of the Marans derives from the wild species of the “Bankiva” type, whose original plumage is Black Red. This “Gilded Salmon" [or Black Red] colouration offers a hen perfectly adapted camouflage enabling her to successfully brood chicks on the ground, as they are exposed to danger.
With domestication appears mutations, making it possible to bring differences to the “wild” variety, for example, such as the Partridge and Wheaten varieties, which are differences relating to only the hen’s plumage, because that of the cock remains almost identical. The homogeneitiy of the wild species’s plumage is less explained by real genetic homogeneity than by the fact that any unfavorable variation is immediately eliminated by nature for the survival of the species (for example: albinos).
In the absence of this pressure of natural selection, genetic modifications are observed with the appearance of new phenotypes. That makes it possible to affirm that the wild species holds in its genetic capital a great deal of diversity which only requires that certain environmental conditions be modified to be expressed.
With regard to plumage, some of these characteristics are dominant, such as Black.
By consanguineous matings, the recessive colours like White or Wheaten, for example, then have a greater chance of becoming homozygous.
In addition, the Marans is as particularly rich in colour varieties as its ancestors. It appears theoretically possible that all six existing genetic families of plumage colour exist in the Marans, which is exceptional among purebred poultry. The genetic transmission of traits in poultry is complex and not yet completely understood but the reasoning is strictly mathematical. On behalf of all stockbreeders, it is essential to respect to the maximum genetic consistency of a given variety and avoid any imagination or whim which would not have been naturally shown by a reliable hereditary reasoning.
With regard to the mission of the Marans-Club de France: it is much more significant today to preserve, seriously and rigorously, the many current varieties and those which are genetically and directly derived from it, likely from homologation (for example, Silvered Black [Birchen], Blue-copper or Silvered-Blue [Blue Birchen], which derives from Black-copper [Brown-Red], rather than to create new ones.
FOOT-NOTE: Even if of ideal type and plumage colour, stock which has lost the characteristic of laying extra-russet-red eggs (as defined by the MCF) will never again be true Marans.
Translated from the French by Katherine Anderson (Marans Club USA).
![]()
©
(copyright) MCF 2001/2017
all rights reserved

